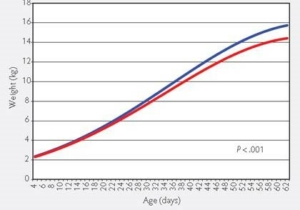
Vitamines als ondersteuning bij varkens; vruchtbaarheid, immuniteit en het bewegingsstelsel
Het is algemeen bekend dat vitaminen belangrijk zijn voor een goede diergezondheid. In het kader van de reductie van het antibioticumgebruik worden aanvullende diervoeders steeds vaker ingezet om problemen te voorkomen. Maar welke vitamines kunnen op welk moment ingezet worden? Om u te ondersteunen bij het correcte gebruik van vitamines wordt in dit artikel beschreven welke vitamines mogelijk een rol spelen bij de vruchtbaarheid, immuniteit en het bewegingsstelsel. Dit is zeker geen volledig overzicht; effecten op productie en effecten op andere factoren worden hier niet besproken. Voor meer informatie over de literatuur betreffende vitaminen en mineralen bij varkens en andere diersoorten verwijzen we u naar dit overzicht.
Vruchtbaarheid
Er zijn verschillende factoren binnen de varkenshouderij die van invloed zijn op de vruchtbaarheid van varkens. De juiste voorziening van vitaminen is hier één van.
Biotine
Een biotine deficiëntie kan een oorzaak zijn van een slechte vruchtbaarheid [1]. Het toedienen van biotine kan leiden tot een toename van de toomgrootte [2, 3] en een toename van het aantal gespeende biggen per zeug [4-6]. In een andere studie wordt een niet significante toename in het aantal gespeende biggen gevonden.
Zeugen die extra biotine krijgen hebben een hoger drachtpercentage bij de eerste berigheid na de partus en een korter interval tussen spenen en eerste berigheid [7]. Ook in een andere studie wordt aangetoond dat het interval tussen spenen en eerste berigheid kan worden verkort door het toedienen van biotine. Dit leidt ook tot een korter interval tussen spenen en drachtigheid en een daling van het aantal dieren dat behandeld moet worden voor anoestrus [8].
Foliumzuur
Foliumzuur is een belangrijk vitamine als het gaat om vruchtbaarheid [4]. Het toedienen van foliumzuur tijdens de dracht gaat gepaard met een consistente toename van de toomgrootte, waarschijnlijk veroorzaakt door een betere overleving van het embryo en de foetus [9-11]. Ook heeft foliumzuur invloed op de melkproductie; de melkproductie kan toenemen, maar de melk kan ook van samenstelling veranderen, waardoor de biggen beter groeien [12].
Vitamine A
De toediening van vitamine A bij jonge zeugen tijdens het spenen en de dracht zorgt voor een verbetering van het aantal geboren en gespeende biggen per toom [13]. Mogelijk wordt dit verklaard door een betere overleving van de embryo’s [4], maar ook de ontwikkeling van de eicellen en embryo’s lijkt beïnvloed te worden door vitamine A. Het aantal eicellen dat in een vergevorderd stadium van de ontwikkeling is neemt toe en de variatie in ontwikkeling neemt af [14]. Ondanks de resultaten die bereikt worden bij een langdurigere toediening, heeft een eenmalige injectie met vitamine A geen effect op de vruchtbaarheid van zeugen [15].
Niet alleen de vruchtbaarheid van zeugen wordt beïnvloed door vitamine A, het is ook belangrijk voor de vruchtbaarheid van beren [16].
B-vitaminen
Ook vitamine B2 is een belangrijk vitamine voor de vruchtbaarheid [1]. Degeneratie van de ovaria kan het gevolg zijn van een deficiëntie [4], maar ook anoestrus, embryonale sterfte en reabsorptie, premature geboorte van biggen, de geboorte van zwakke biggen, en een hoge mortaliteit van biggen in de eerste twee levensdagen [17, 18] kunnen voorkomen bij een tekort aan vitamine B2. Andere studies laten ook een verband zien tussen de toomgrootte en mortaliteit van de biggen [18-20].
Een vitamine B5 deficiëntie kan leiden tot atrofie van de ovaria en een daling van de oestrogeen en progesteron productie [1].
Vitamine B6 speelt ook een rol bij de vruchtbaarheid; het toedienen van vitamine B6 aan zeugen met een deficiëntie zorgt voor een toename van de toomgrootte [4].
Vitamine E
Bij zeugen lijkt een vitamine E deficiëntie een risicofactor te zijn voor een verhoogde incidentie van het MMA- syndroom [4, 21]. Ook is er een verband aangetoond tussen de vitamine E voorziening en de toomgrootte. De toediening van vitamine E leidt tot een toename van het aantal geboren biggen [21] en het aantal levende biggen op een leeftijd van zeven dagen [22].
Ook blijkt de toediening van vitamine E aan zeugen tijdens dracht en lactatie belangrijk voor de vitamine E voorziening van de biggen na de partus [4, 21-23].
Hoewel vitamine E altijd in het voer zit, kunnen deficiënties toch voorkomen. De vitamine E behoefte van dieren is namelijk afhankelijk van verschillende factoren zoals de seleniumvoorziening en de hoeveelheid antioxidanten in het voer. Bovendien is de voorziening via het voer niet altijd betrouwbaar, omdat vitamine E snel degradeert onder invloed van warmte, vocht, vet of spoorelementen [4].
Immuniteit
Biotine
Biotine is belangrijk voor een goede werking van het immuunsysteem van varkens [24]. Een extreem hoge toediening van biotine (>880 mg/kg voer) kan echter juist zorgen voor remming van het immuunsysteem [24].
Vitamine A
Een suboptimale vitamine A voorziening leidt tot een daling van de antilichaam- en lymfocytenproductie [1]. Een deficiëntie bij zeugen kan leiden tot een vitamine A deficiëntie bij de geboren biggen. Dit zorgt bij deze biggen voor een verminderde immuunreactie na blootstelling aan het rotavirus [25, 26].
B-vitaminen
Voor vitamine B2 en B5 is weinig bekend over de relatie tot het immuunsysteem. Wel is bekend dat een deficiëntie kan zorgen voor een slechte immuunreactie [4].
Vitamine C
Vitamine C is van belang voor de fagocytotische activiteit van leukocyten en de productie van antilichamen. Daarnaast is vitamine C belangrijk door zijn werking als antioxidant [1, 4, 27].
Het toedienen van extra vitamine C aan varkens kan zorgen voor een toename in het aantal leukocyten, een verbeterde levensduur van leukocyten en een verbeterde fagocytotische activiteit van leukocyten in het bloed en in de longen [28].
Vitamine D
In één studie is aangetoond dat het toedienen van vitamine D tijdens een challenge met rotavirus zorgt voor een vermindering van de negatieve effecten op het lichaamsgewicht, de voeropname, de villuslengte en consistentie van de mest [29].
Vitamine E
Een vitamine E deficiëntie heeft ook invloed op het immuunsysteem, waarschijnlijk door remming van de lymfocytenproductie [30] en de activiteit van lymfocyten en polymorf nucleaire leukocyten [31]. De productie van antilichamen kan toenemen door de toediening van vitamine E [32-34]. Het effect van vitamine E op het immuunsysteem van biggen wordt ook bereikt wanneer vitamine E aan de zeugen wordt toegediend [23].
Effecten op het immuunsysteem na de toediening van vitamine E zijn niet alleen meetbaar aan de hand van bloedwaarden, maar leiden ook tot verbetering op het bedrijf. Op één bedrijf verminderde de ernst van diarree en werd de mortaliteit verlaagd [35]. Op een ander bedrijf zorgde de toediening van vitamine E voor een dusdanige vermindering van problemen met speendiarree dat minder antibiotica nodig waren [36]. PRRS problemen konden op een ander bedrijf echter niet worden verminderd door het gebruik van vitamine E [37].
Het bewegingsstelsel
Biotine
Biotine wordt in het maagdarmkanaal van varkens geproduceerd, maar toch kan een deficiëntie optreden [38].
Een biotine deficiëntie kan onder andere leiden tot afwijkingen aan de huid en het hoorn van de hoeven [1, 38], met zwak, broos en soms zelfs necrotisch hoornweefsel tot gevolg [39]. Door het toedienen van extra biotine kunnen de hoorndichtheid [4, 40] en de drukweerstand van het hoorn verbeteren [4, 41]. Ook het aantal scheuren in hoeven, witte lijn defecten en voetzoollaesies kunnen afnemen wanneer extra biotine wordt gegeven [4, 42, 43].
Uit een Nederlandse studie is gebleken dat het consequent toedienen van biotine aan zeugen kan zorgen voor een vermindering van het aantal zeugen dat afgevoerd moet worden ten gevolge van kreupelheid van 25% naar 14%. Het aantal laesies bij gelten daalde met 52% [44]. In een andere studie zijn ook positieve effecten gevonden van het toedienen van biotine op de hoefgezondheid, maar hier daalde de afvoer van zeugen niet [45].
Het effect dat gezien wordt na de toediening van biotine is afhankelijk van de dosering; hoe hoger de dosering, hoe sneller een positief effect wordt waargenomen op de incidentie van hoefafwijkingen [46].
Vitamine C
Vitamine C is ook belangrijk voor het bewegingsapparaat, omdat het belangrijk is voor de productie van collageen in kraakbeen en botweefsel [1, 4]. Een vitamine C deficiëntie bij zeugen kan leiden tot osteochondrose [4].
Vitamine D
Een vitamine waarvan algemeen bekend is dat het belangrijk is voor het bewegingsapparaat is vitamine D. Vitamine D speelt een rol in de absorptie en verwerking van calcium en fosfor. Een deficiëntie leidt tot rachitis bij biggen en osteomalacie bij volwassen dieren [1, 4, 47]. De toediening van extra vitamine D kan zorgen voor een normale endochondrale ossificatie, een remming van osteochondrose en een toename van de regeneratie van beschadigd kraakbeen [48].
Vitamine E
Tot slot is ook vitamine E belangrijk voor het bewegingsapparaat. Een deficiëntie kan namelijk, naast andere verschijnselen, leiden tot degeneratie van spieren en botweefsel [1, 4].
Dopharma
Dopharma heeft een aanvullend diervoeder met vitaminen in het assortiment:
- Osteosol AD bevat enkele vitaminen, maar daarnaast ook mineralen/spoorelementen.
Hiernaast hebben we ook drie diergeneesmiddelen in het assortiment die vitaminen bevatten:
- Vitasol multi is een vloeibaar product met vitamines.
- Vitaminsol multi is een poedervormig product met een combinatie van vitamines en mineralen/spoorelementen.
- Vitasol® C is een geregistreerd diergeneesmiddel met vitamine C.
Referenties
- Isabel, B. and A.I.B. Rey, L.C., Optimum vitamin nutrition in pigs, in Optimum vitamin nutrition; in the production of quality animal foods. 2012, 5M Publishing: United Kingdom. p. 243-306.
- Penny, R.H., et al., Influence of biotin supplementation on sow reproductive efficiency [Abstract]. Vet Rec, 1981. 109(4): p. 80-1.
- Brooks, P.H., D.A. Smith, and V.C. Irwin, Biotin-supplementation of diets; the incidence of foot lesions, and the reproductive performance of sows [Abstract]. Vet Rec, 1977. 101(3): p. 46-50.
- NRC, Nutrient requirement of swine. 10th ed. 1998, Washington DC: National Academy of Science, National Research Council.
- Lewis, A.J., G.L. Cromwell, and J.E. Pettigrew, Effects of supplemental biotin during gestation and lactation on reproductive performance of sows: a cooperative study. J Anim Sci, 1991. 69(1): p. 207-14.
- Hamilton, C.R. and T.L. Veum, Response of sows and litters to added dietary biotin in environmentally regulated facilities [Abstract]. J Anim Sci, 1984. 59(1): p. 151-7.
- Bryant, K.L., et al., Supplemental biotin for swine. II. Influence of supplementation to corn- and wheat-based diets on reproductive performance and various biochemical criteria of sows during four parities [Abstract]. J Anim Sci, 1985. 60(1): p. 145-53.
- Simmins, P.H. and P.H. Brooks, Supplementary biotin for sows: effect on reproductive characteristics [Abstract]. Vet Rec, 1983. 112(18): p. 425-9.
- Lindemann, M.D., Supplemental folic acid: a requirement for optimizing swine reproduction. J Anim Sci, 1993. 71(1): p. 239-46.
- Thaler, R.C., et al., Effect of dietary folic acid supplementation on sow performance through two parities [Abstract]. J Anim Sci, 1989. 67(12): p. 3360-9.
- Lindemann, M.D. and E.T. Kornegay, Folic acid supplementation to diets of gestating-lactating swine over multiple parities [Abstract]. J Anim Sci, 1989. 67(2): p. 459-64.
- Wang, S.P., et al., Effects of folic acid on the performance of suckling piglets and sows during lactation [Abstract]. J Sci Food Agric, 2011. 91(13): p. 2371-7.
- Lindemann, M.D., et al., A regional evaluation of injections of high levels of vitamin A on reproductive performance of sows. J Anim Sci, 2008. 86(2): p. 333-8.
- Whaley, S.L., et al., Influence of vitamin A injection before mating on oocyte development, follicular hormones, and ovulation in gilts fed high-energy diets. J Anim Sci, 2000. 78(6): p. 1598-607.
- Pusateri, A.E., M.A. Diekman, and W.L. Singleton, Failure of vitamin A to increase litter size in sows receiving injections at various stages of gestation. J Anim Sci, 1999. 77(6): p. 1532-5.
- Booth, W.D., Vitamin A in testicular tissue of the boar and intersex pig. J Reprod Fertil, 1974. 40(1): p. 219-22.
- Esch, M.W., R.A. Easter, and J.M. Bahr, Effect of riboflavin deficiency on estrous cyclicity in pigs. Biol Reprod, 1981. 25(3): p. 659-65.
- Pettigrew, J.E., et al., Riboflavin nutrition of sows. J Anim Sci, 1996. 74(9): p. 2226-30.
- Bazer, F.W. and M.T. Zavy, Supplemental riboflavin and reproductive performance of gilts. Journal of Animal Science, 1988. 66, suppl 1: p. 324.
- Frank, G.R., J.M. Bahr, and R.A. Easter, Riboflavin requirement of gestating swine. J Anim Sci, 1984. 59(6): p. 1567-72.
- Mahan, D.C., Effects of dietary vitamin E on sow reproductive performance over a five-parity period. J Anim Sci, 1994. 72(11): p. 2870-9.
- Mahan, D.C., Assessment of the influence of dietary vitamin E on sows and offspring in three parities: reproductive performance, tissue tocopherol, and effects on progeny. J Anim Sci, 1991.69(7): p. 2904-17.
- Babinszky, L., et al., Effect of vitamin E and fat source in sows’ diets on immune response of suckling and weaned piglets. J Anim Sci, 1991. 69(5): p. 1833-42.
- Kornegay, E.T., et al., Effects of biotin and high copper levels on performance and immune response of weanling pigs. J Anim Sci, 1989. 67(6): p. 1471-7.
- Eriksson, M., et al., Beneficial effects of pre-treatment with vitamin A on cardiac and pulmonary functions in endotoxaemic pigs. Acta Anaesthesiol Scand, 1996. 40(5): p. 538-48.
- Eriksson, M., et al., Vitamin A exerts potential therapeutic effects in the endotoxaemic pig. Acta Anaesthesiol Scand, 1997. 41(7): p. 824-9.
- Zhao, J., et al., Effects of vitamin C supplementation on performance, iron status and immune function of weaned piglets. Arch Tierernahr, 2002. 56(1): p. 33-40.
- Konowalchuk, J.D., et al., Modulation of weanling pig cellular immunity in response to diet supplementation with 25-hydroxyvitamin D(3). Vet Immunol Immunopathol, 2013. 155(1-2): p. 57-66.
- Zhao, Y., et al., Dietary vitamin D supplementation attenuates immune responses of pigs challenged with rotavirus potentially through the retinoic acid-inducible gene I signalling pathway [Abstract]. Br J Nutr, 2014. 112(3): p. 381-9.
- Lessard, M., et al., Cellular immune responses in pigs fed a vitamin E- and selenium-deficient diet. J Anim Sci, 1991. 69(4): p. 1575-82.
- Wuryastuti, H., et al., Effects of vitamin E and selenium on immune responses of peripheral blood, colostrum, and milk leukocytes of sows. J Anim Sci, 1993. 71(9): p. 2464-72.
- Ellis, R.P. and M.W. Vorhies, Effect of supplemental dietary vitamin E on the serologic response of swine to an Escherichia coli bacterin. J Am Vet Med Assoc, 1976. 168(3): p. 231-2.
- Peplowski, M.A., et al., Effect of dietary and injectable vitamin E and selenium in weanling swine antigenically challenged with sheep red blood cells. J Anim Sci, 1980. 51(2): p. 344-51.
- Hayek, M.G., et al., Porcine immunoglobulin transfer after prepartum treatment with selenium or vitamin E [Abstract]. J Anim Sci, 1989. 67(5): p. 1299-306.
- Agger, N., L. Pontoppidan, and R.L. Stuart, Increased Immunity in Weaned Piglets by Daily Dosing of d-alpha-Tocopherol in the Drinking Water. Acta Veterinaria Scandinavica 2003. 44 (Suppl 1): p. 108.
- Lamberts, F.J., Vitamin E as a possible aid in the control of disease problems on pig farms: a field test. Tijdschr Diergeneeskd, 1997. 122(7): p. 190-2.
- Toepfer-Berg, T.L., et al., Vitamin E supplementation does not mitigate the acute morbidity effects of porcine reproductive and respiratory syndrome virus in nursery pigs. J Anim Sci, 2004.82(7): p. 1942-51.
- Kornegay, E.T., Biotin in swine nutrition. Ann N Y Acad Sci, 1985. 447: p. 112-21.
- Fritsche, A., G.A. Mathis, and F.R. Althaus, Pharmacologic effects of biotin on epidermal cells. Schweiz Arch Tierheilkd, 1991. 133(6): p. 277-83.
- Kempson, S.A., R.J. Currie, and A.M. Johnston, Influence of biotin supplementation on pig claw horn: a scanning electron microscopic study. Vet Rec, 1989. 124(2): p. 37-40.
- Webb, N.G., R.H. Penny, and A.M. Johnston, Effect of a dietary supplement of biotin on pig hoof horn strength and hardness. Vet Rec, 1984. 114(8): p. 185-9.
- Penny, R.H., et al., Foot rot of pigs: the influence of biotin supplementation on foot lesions in sows [Abstract]. Vet Rec, 1980. 107(15): p. 350-1.
- Bryant, K.L., et al., Supplemental biotin for swine. III. Influence of supplementation to corn- and wheat-based diets on the incidence and severity of toe lesions, hair and skin characteristics and structural soundness of sows housed in confinement during four parities [Abstract]. J Anim Sci, 1985. 60(1): p. 154-62.
- de Jong, M.F. and J.R. Sytsema, Field experience with d-biotin supplementation to gilt and sow feeds [Abstract]. Vet Q, 1983. 5(2): p. 58-67.
- Simmins, P.H. and P.H. Brooks, Supplementary biotin for sows: effect on claw integrity [Abstract]. Vet Rec, 1988. 122(18): p. 431-5.
- Misir, R. and R. Blair, Effect of biotin supplementation of a barley-wheat diet on restoration of healthy feet, legs and skin of biotin deficient sows [Abstract]. Res Vet Sci, 1986. 40(2): p. 212-8.
- Daculsi, G., B. Kerebel, and L.M. Kerebel, Effect of vitamin D3 and vitamin D3 sulfate on dental and bone tissues in the pig. J Biol Buccale, 1981. 9(4): p. 363-74.
- Sugiyama, T., et al., Effects of 25-hydroxy-cholecalciferol on the development of osteochondrosis in swine. Anim Sci J, 2013. 84(4): p. 341-9.